Carotenoids, retinoids modulate ovarian development in Litopenaeus vannamei
Can protect reproductive tissue from toxic levels of free oxygen radicals

Functional metabolites like caro-tenoids are bioactive ingredients with physiological or health benefits beyond basic nutritional functions. The levels of carotenoids in commercial shrimp feeds are generally insufficient under intensive culture conditions to reach the physiological levels needed for critical bioactive functions in shrimp. As a result, the animals may lose their natural capacity for metabolic bioconversion to more bioactive and functional forms, such as retinoids.
The consequences include a decrease in their immunoresistance, development of diseases, and interruption of critical processes like maturation and reproduction. A study evaluated the bioactive nonpigmenting functions of carotenoids and retinoids in reproductive organs during maturation of the white shrimp (Litopenaeus vannamei).
Carotenoids and retinoids
Carotenoids are nonproduced de novo metabolites and natural components in the diet of shrimp and many other marine organisms. Once ingested by shrimp, carotenoid molecules are unstable and chemically reversible.
Retinoids are a class of molecules derived from vitamin A considered crucial for many growth, development, and homeostatic processes in animals. They are considered to be hormone-like compounds. However, the functions of retinoids in penaeid shrimp have not been sufficiently studied.
According to the metabolic pathway of crustaceans, carotenoids can derive into other metabolites such as retinoids. The study determined that carotenoids are natural precursors of retinoids in L. vannamei.
Study setup
In this study, different concentrations of beta-carotene (18.6 μg per gram), astaxanthin (18.6 μg per gram), retinal palmitate (5.3, 10.6, 16.0 μg per gram) and all-trans retinoic acid (133, 266, 400 μg per gram) were administered by injection to farmed, reproductively exhausted female L. vannamei females in gonadic conditions II and IV, respectively. The shrimp had been formerly eyestalk ablated.
Results
After the experimental period, carotenoids, retinoids, and their respective derived metabolites were analyzed. Immunohistochemistry was also performed to detect receptors of retinoids. Table 1 shows the effects of the carotenoids and retinoids administered.
Paniagua-Michel, Survival and average value of gonodosomatic (GSI) and hepatosomatic (HSI) indexes, Table 1
Table 1. Survival and average value of gonodosomatic (GSI) and hepatosomatic (HSI) indexes in L. vannamei broodstock under experimental conditions (n = 13).
It was determined that retinoic acid had a teratogenic effect in the shrimp. Results showed the treatment based on retinol palmitate significantly enhanced (p < 0.05) gonadic maturation and survival in first term, with 40 percent of the population reaching maturation by day 5 of treatment. The study also showed that astaxanthin and beta-carotene play an important role not only in modulating maturation, but also as precursors of retinoids at different levels of bioactivity.
New metabolites
The first registered presence in L. vannamei of isomers 13-cis retinol, 9-cis retinol, all-trans retinal, and all-trans retinol indicated functional roles as effectors of maturation and reproduction. Results indicated the bioconversion of carotenoids into retinoids occurs in L. vannamei, and that both oxygenic carotenoids (mainly astaxanthin and others involved in shrimp metabolic pathways) and anoxygenic carotenoids (mainly beta-carotene) must be supplied at physiological levels in the diet to achieve pigmentation and bioactive functions such as full conversion to retinoids.
Proposed model
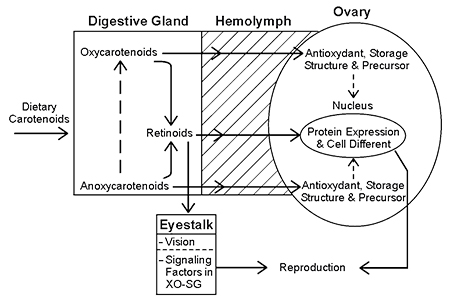
Fig. 1 shows the proposed model of bioconversion of dietary carotenoids into retinoids in the organs involved in the maturation of penaeid shrimp. Different carotenoids show different degrees of bioconversion to retinoids in shrimp. Oxygenic carotenoids exhibit less bioconversion properties, while anoxygenic carotenoids are readily bioconverted to two molecules of retinoids. The uptake of dietary carotenoids is bioconverted to retinoids in the digestive gland and carried in carotenolipoproteins in the haemolymph to the gonads, where more specialized retinoids are produced.
In some crustaceans and other arthropods, isomers of retinoids bind to hormonal nuclear receptors. These receptors are known to play important functions as signaling factors for protein and cellular differentiation.
Conclusion
Results of this study reinforced the need for inclusion of carotenoids in commercial shrimp diets and showed that in L. vannamei, retinoid metabolites induce growth and development by controlling the production of local morphogenic signals by a nuclear receptor signaling pathway, which enhances maturation and associated processes. At this level, carotenoids play a role in protecting reproductive tissue from toxic levels of free oxygen radicals, and as a reserve during embryogenesis and development.
(Editor’s Note: This article was originally published in the October 2002 print edition of the Global Aquaculture Advocate.)
Now that you've finished reading the article ...
… we hope you’ll consider supporting our mission to document the evolution of the global aquaculture industry and share our vast network of contributors’ expansive knowledge every week.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year. GSA individual and corporate members receive complimentary access to a series of GOAL virtual events beginning in April. Join now.
Not a GSA member? Join us.
Authors
-

Centro de Investigación Científica y de Educación Superior de Ensenada
Department of Marine Biotechnology
Km 107 Carretera Tijuana-Ensenada
Ensenada, BC Mexico -
Centro de Investigación Científica y de Educación Superior de Ensenada
Department of Marine Biotechnology
Km 107 Carretera Tijuana-Ensenada
Ensenada, BC Mexico
Related Posts

Aquafeeds
Astaxanthin: Feed supplement enhances pigmentation, health
Dietary carotenoids maintain optimal pigmentation and health benefits in cultured aquatic animals. Researchers consider astaxanthin an essential nutrient.

Health & Welfare
Canthaxanthin stands up to scrutiny
Carotenoids like canthaxanthin are biological pigments widely found in nature that provide color to many species of birds, insects, crustaceans and fish.

Aquafeeds
Microalgae provide food, feed, fuel
About 30 percent of global microalgae production goes to animal feeds, with large applications in aquaculture, particularly for larval stages.

Aquafeeds
Palm oil: Alternative lipid source in aquaculture feeds
One potential substitute for fish oil in aquafeeds is palm oil, which is the second most abundant vegetable oil in the world.