Evaluating temperature preference of Nile tilapia juveniles
Results shows species can briefly choose a masculinizing temperature

The main environmental sex determination factor reported in fish is temperature, and almost 60 fish species displaying temperature sex determination or genetic sex determination (GSD) with a temperature effect (GSD+TE) have been described in the literature.
Nile tilapia (Oreochromis niloticus) is an African freshwater fish living in contrasting habitats. Thermal treatments with temperatures above 32 degrees-C to 36.5 degrees-C for at least 10 days during the gonadal differentiation period [from 10 to 30 days post fertilization (dpf) for Nile tilapia] induce masculinization. The magnitude of masculinization strongly depends on parental effects, from no effect to almost 100 percent males.
In Nile tilapia, the optimal growing temperature in a controlled environment is approximately 27 to 30 degrees-C, below the masculinizing high temperatures above 32 degrees-C. Thus, the relevance of thermal sex reversal in a natural context and the consequences for potential selective advantages are unclear. Additionally, it was unclear whether undifferentiated O. niloticus juveniles would spontaneously move to high, potentially masculinizing temperatures, inducing sex reversal. The thermal preference of species exhibiting a GSD + TE sex determining system during the thermosensitive period for sex determination has only been studied in African catfish.
This article – adapted and summarized from the original publication – reports on a study to determine whether sexually undifferentiated juveniles of Nile tilapia spontaneously prefer higher, unfavorable temperatures and whether this choice skews the sex ratio toward males.
Study setup
Nile tilapias from the Lake Manzala (Egypt) strain were obtained from the Research and Education Center in Aquaculture (CEFRA), Liège University, Belgium. All matings were performed between XX females and XX males, the latter obtained by hormonal sex reversal. XX progenies were used to better detect a potential masculinizing effect.
Five full-sib progenies (from 100 percent XX crosses) were subjected to (1) a horizontal three-compartment thermal step gradient (thermal continuum 28 to 32 to 36.5 degrees-C) during the thermosensitive period, (2) a control continuum (28- 28- 28 degrees-C) and (3) a thermal control tank (36.5 degrees-C).
For detailed information on the experimental setup; progenies; rearing conditions; continuum compartmental occupation analysis; survival and sex ratio analyses; statistical analysis of data and other experimental design aspects, please refer to the original publication.
Results and discussion
Nile tilapia have a complex sex determination mechanism with major and minor genetic factors and a temperature effect (GSD+TE). Our experiment was designed to provide some insight into the relevance of temperature effects on Nile tilapia sex determination when fish have the opportunity to choose their thermal preference in a horizontal thermal step gradient, which represents some temperature choices that fish could make at a specific period of their life in the wild.
Our study shows that sexually undifferentiated juvenile tilapia moved across a three-connected-compartment device (continuum). In a control continuum (CC) where temperatures in the three compartments were identically set to 28 degrees-C at all times, the 10 dpf juveniles evenly occupied the three compartments after the first 5 days and that distribution stayed essentially even until the end of the experiment (40 dpf). Similar results have been reported by other researchers and other fish species.
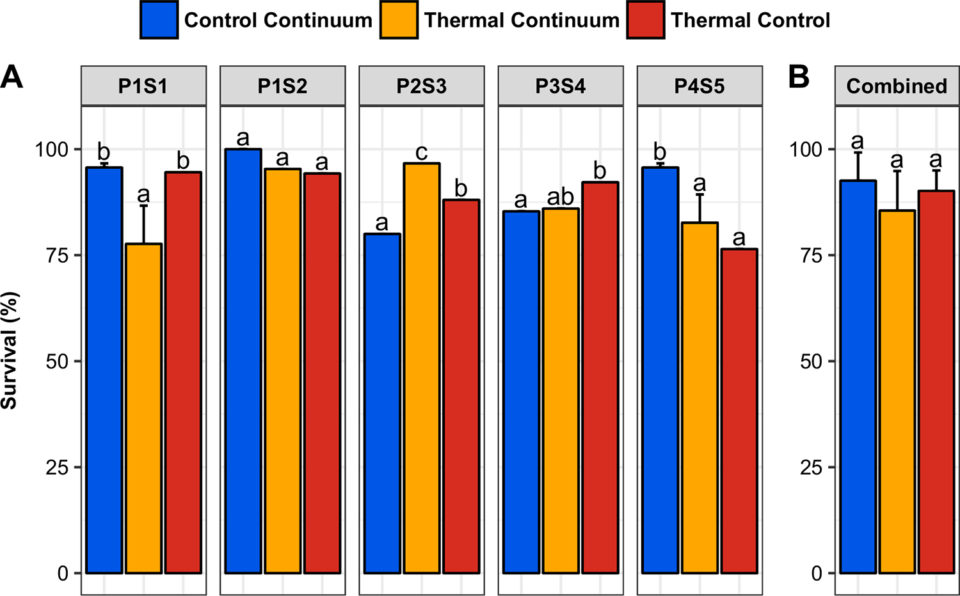
Survival rates at 40 dpf (post-treatment) in the control continuum, thermal continuum and thermal control (A: by progeny; B: mean + SD). Within each chart, bars with the same letter are not significantly different (generalized linear model, p-value > 0.05).
Animal personality traits also influence behavior in a context-dependent manner. In fish, three personality types have been described: 1) proactive: active coping, risk takers, suggesting a higher energy requirement, 2) reactive: passive coping, shy with a low level of aggression and immobility, suggesting lower metabolic costs, and 3) intermediate individuals between these two extremes.
In Nile tilapia and zebra fish, proactive individuals exhibit a higher thermal preference compared to reactive individuals. In our experiments, we used unsorted batches; thus, the distribution of thermal preferendum observed should reflect a mixed personality population. Interestingly, such an interpretation would lead us to conclude that the individuals preferring the higher temperatures, in particular those that prefer 36.5 degrees-C during the initial period and later on, would be the most proactive, exploratory individuals.
Existence of two successive thermal preferenda in our experiment could be related to the age and size of the fish. The optimal temperature for growth evolves depending on body mass in Oreochromis aureus, a species closely related to O. niloticus. Further experiments will be required to clearly address this issue.
It was previously stated that temperatures above 32 to 35 degrees-C can induce masculinization in Nile tilapia. Our results, together with those of other researchers, suggest that high temperature masculinization could take place inside a thermal gradient during a very short period shortly after 10 dpf. Interestingly, this time point corresponds to the end of the strictly maternal mouth-brooding period, when juveniles are released and can experiment potentially masculinizing temperatures in the shallow margins of water bodies.
In the present study, we observed that diffusion kinetics in the thermal continuum varied across progenies. This suggests that the more proactive, exploratory populations may be more prone to exposure to higher temperatures (possibly also exploring compartment 3 more intensely, see above) and would thus experience the highest sex reversal rates.
The overarching question concerning temperature-induced (or, in general, environmentally induced) sex reversal is whether this phenomenon is actively maintained through evolution in certain species to better cope with potentially dangerous situations or to adapt the sex ratio in changing environments. What could be the selective advantage of sex reversal for the survival of a species?
Our study shows that, when presented with a choice of different thermal conditions during the sex determination period, Nile tilapia juveniles tend to prefer higher temperatures even though they experience masculinization. This indicates that such a behavior is at least not deleterious to the population, otherwise, it would have been avoided. Whether this behavior is actually beneficial for the population remains to be determined. It is interesting to note that our observations show that there is a familial component in the extent of sex reversal observed, as well as a clear link with the exploratory character of a population.
Perspectives
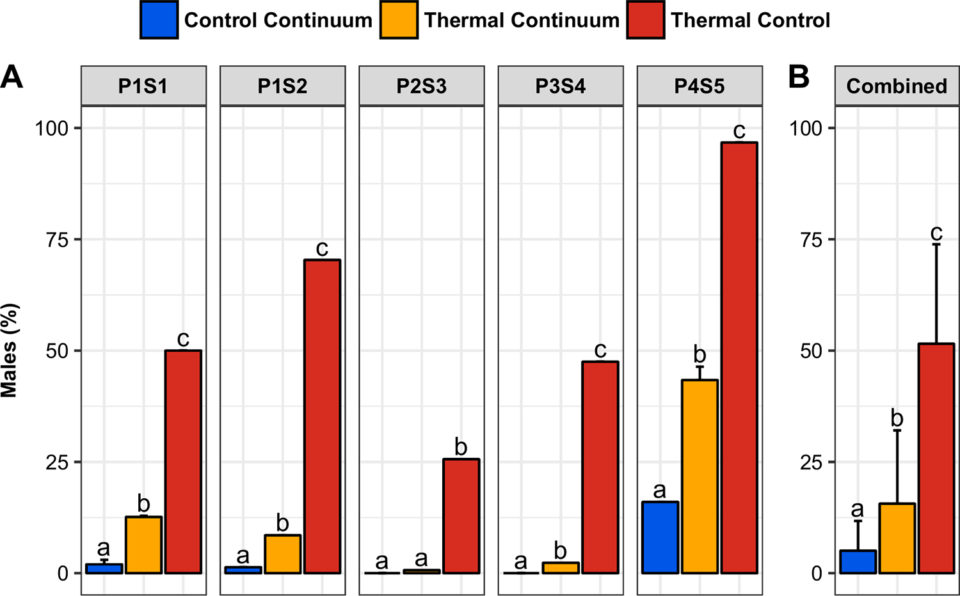
In our study, during the first days of the treatment, up to an average of 20 percent of the population preferred the masculinizing compartment of the thermal continuum (36.5 degrees-C) compared to the control continuum. During the second part of the treatment, juveniles preferred the lower, non-masculinizing 32 degrees-C temperature. This short exposure to higher temperatures was sufficient to significantly skew the sex ratio toward males, compared to congeners raised at 28 degrees-C (from 5.0 ± 6.7 percent to 15.6 ± 16.5 percent of males).
The proportion of males was significantly different in the thermal continuum, thermal control tank and control continuum, and it was positively correlated among populations. Our study shows for the first time that Nile tilapia juveniles can choose a masculinizing temperature during a short period of time. This preference is sufficient to induce sex reversal to males within a population. For the first time, behavior is reported as a potential player in the sex determination mechanism of this species.
Our data also shows that fish migration in a horizontal thermal step gradient depends on multiple factors and evolves during the course of the day. In a thermal continuum, undifferentiated juveniles (1) spontaneously migrate to the higher, potentially masculinizing temperatures during the critical period of sex differentiation; and (2) this migration to high temperatures is sufficient to induce variable but significantly skewed sex ratios toward males. The higher the intrinsic thermo-sensitivity and ability to generate spontaneous sex-reversed males are, the higher the male proportion in the thermal continuum is.
Finally, the factors that motivate this preference for high masculinizing temperatures remain to be elucidated.
Now that you've finished reading the article ...
… we hope you’ll consider supporting our mission to document the evolution of the global aquaculture industry and share our vast network of contributors’ expansive knowledge every week.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year. GSA individual and corporate members receive complimentary access to a series of GOAL virtual events beginning in April. Join now.
Not a GSA member? Join us.
Authors
-

Research and Education Center in Aquaculture (CEFRA), Liège University, Tihange, Belgium, Laboratory for Organogenesis and Regeneration (LOR), Interdisciplinary research institute in the biomedical sciences (GIGA-I3), Liège University, Sart Tilman, Liège, Belgium
-
Research and Education Center in Aquaculture (CEFRA)
Liège University, Tihange, Belgium -
Research and Education Center in Aquaculture (CEFRA)
Liège University, Tihange, Belgium -
Laboratory for Organogenesis and Regeneration (LOR)
Interdisciplinary research institute in the biomedical sciences (GIGA-I3)
Liège University, Sart Tilman, Liège, Belgium -
Laboratory for Organogenesis and Regeneration (LOR)
Interdisciplinary research institute in the biomedical sciences (GIGA-I3)
Liège University, Sart Tilman, Liège, Belgium -
Research and Education Center in Aquaculture (CEFRA)
Liège University, Tihange, Belgium -
Research and Education Center in Aquaculture (CEFRA)
Liège University, Tihange, Belgium
Related Posts

Health & Welfare
Blue catfish outproduce channel catfish under low-D.O. conditions
Although there is increasing interest in blue catfish, a potential disadvantage of the fish when compared to channel catfish is their reported poorer tolerance of low dissolved-oxygen concentrations.

Health & Welfare
Artificial incubation, hormonal sex reversal promoted tilapia boom
The tilapia production boom results from technology that combines artificial egg incubation and monosex fry production using hormonal sex reversal.

Health & Welfare
GIFT tilapia show greater FCR, growth potential than red tilapia
In a study of the effects of interaction between genotype and dietary protein levels on fish performance, GIFT tilapia had greater growth than red hybrids fed diets with equal or higher protein levels.

Health & Welfare
China’s tilapia germplasm: Chance and challenge
China’s expansive tilapia culture industry is based on exotic species introduced continuously since 1956. Technical advances for improving the limited germplasm are expected to aid the industry.