Kona-line SPF shrimp, Vibrio penaeicida candidates for bacterial challenge model
Direct relationship observed between bacterial concentrations and shrimp mortality

Most shrimp disease challenge models have been developed and standardized for the study of diseases caused by such specific viruses as White Spot Syndrome Virus and Taura Syndrome Virus. These challenge models are an important component of shrimp-breeding programs as an aid in disease-resistance selection. They are also used to evaluate the effects of environmental factors on the manifestation of the diseases and in the evaluation of possible therapeutic and preventative agents.
Bacterial challenge models, in comparison, are less developed. Although very important, the economic impacts of bacterial diseases on the shrimp-farming industry are apparently not as severe as those of some viral diseases, so less interest and research funding are channeled in that direction.
Additionally, bacterial challenge models are very difficult to standardize because shrimp’s responses to infection are highly dependent on their physiological condition, the source of the shrimp, bacterial species and strains, route of infection, environmental conditions, and other factors. Moreover, some of the most common bacteria pathogenic to shrimp – usually Vibrio species – are opportunistic and the cause of secondary infections.
For challenge studies where healthy shrimp are employed, unusually high numbers of these bacteria are necessary to cause disease. However, such high numbers of bacteria also tend to quickly overwhelm the shrimp’s immune systems, and the shrimp succumb to the disease before any additional testing of therapeutics can be done.
Challenge model
The Kona-line shrimp – specific pathogen-free (SPF) Litopenaeus vannamei shrimp – and the bacteria Vibrio penaeicidan are promising candidates for the development of a bacterial challenge model. An established breeding line of shrimp, such as the SPF Kona line, ensures little or no genetic variation from one experiment to the next. Additionally, the Kona shrimp are commercially available, thus facilitating the reproducibility of experiments.
V. penaeicida are highly pathogenic to penaeid shrimp. They cause severe infection, produce distinctive histological lesions and are relatively easy to cultivate in vitro. The pathogenicity of V. penaeicida has also been studied in L. stylirostris, Penaeus monodon and Marsupenaeus japonicus.
Virulence study
The authors recently conducted a series of three consecutive experiments on virulence assessment where juvenile SPF L. vannamei from the Kona line were challenged by intramuscular injection with varied concentrations of V. penaeicida.
In the first experiment, four groups of 20, 4-gram shrimp received approximately 500,000; 5,000; 500; and 50 colony-forming units (colony-forming unit) bacteria per shrimp, respectively. In the second experiment, four groups of 20 shrimp received approximately 10,000; 5,000; 2,500; and 500 colony-forming unit per shrimp. In the third experiment, four groups of shrimp received approximately 1,000; 500; 250; and 125 colony-forming unit per shrimp.
The two-layer total plate count method was used to estimate bacteria numbers in the concentrated cultures and inocula. A negative control that consisted of 20 shrimp injected with the same sterile 2.5 percent saline used to prepare the inocula was included in each experiment.
The shrimp were maintained in 90-liter tanks at 25-ppt salinity and 27 to 28 degrees-C. Each tank was fitted with its own biological filter. Injected shrimp were monitored every six hours for a maximum of 10 days. Dead shrimp were counted, weighed and discarded. Moribund shrimp were fixed and examined by conventional histology and embryology techniques to confirm the presence of lesions diagnostic of systemic bacterial infection by V. penaeicida.
Results
A direct relationship was observed between the bacterial concentrations and shrimp mortality (Fig. 1). At high concentrations, the mortalities were 100 percent for 500,000 colony-forming unit per shrimp, 95 percent for 10,000 colony-forming unit per shrimp, and 90 percent and 91 percent for the two experiments that included 5,000 colony-forming unit per shrimp.
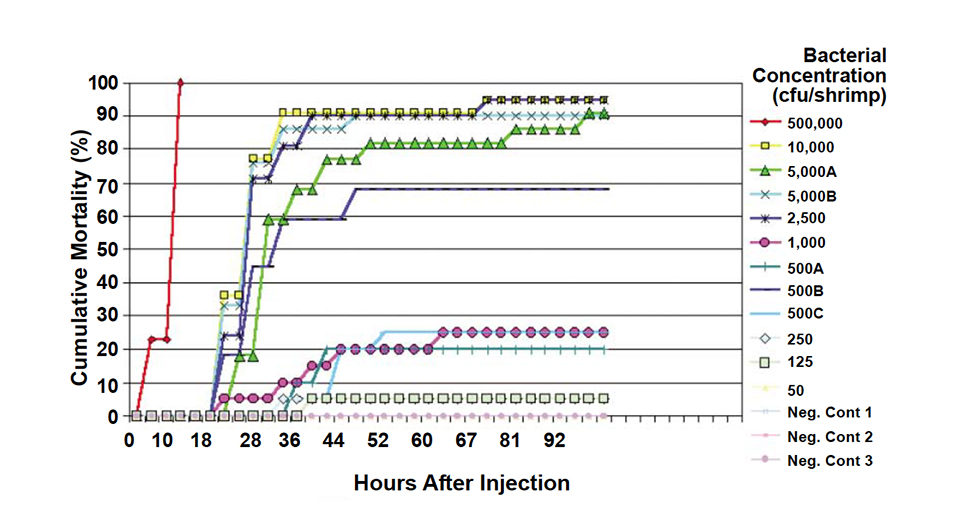
At lower concentrations, the mortalities were 95 percent for 2,500 colony-forming unit per shrimp, 25 percent for 1,000 colony-forming unit per shrimp and 25, 20 and 68 percent for the three experiments that included 500 colony-forming unit per shrimp. Shrimp injected with 250 and 125 colony-forming unit per shrimp showed 5 percent mortality, while those at 50 colony-forming unit per shrimp had no mortality.
Similar percentage mortalities have been reported in L. stylirostris at concentrations within the 200 to 1,000 colony-forming unit per shrimp range.
The experiments indicated that, while causing significant disease, bacterial concentrations of 500 to 5,000 colony-forming unit per shrimp could allow the shrimp to survive long enough for additional testing. Juvenile Kona L. vannamei appear to be an adequate candidate for a challenge model due to their high susceptibility to infection with V. penaeicida.
(Editor’s Note: This article was originally published in the June 2005 print edition of the Global Aquaculture Advocate.)
Now that you've finished reading the article ...
… we hope you’ll consider supporting our mission to document the evolution of the global aquaculture industry and share our vast network of contributors’ expansive knowledge every week.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year. GSA individual and corporate members receive complimentary access to a series of GOAL virtual events beginning in April. Join now.
Not a GSA member? Join us.
Authors
-

Aquaculture Pathology Laboratory
University of Arizona
1117 East Lowell Street
Building 90, Room 106
Tucson, Arizona 85721 USA -
Aquaculture Pathology Laboratory
University of Arizona
1117 East Lowell Street
Building 90, Room 106
Tucson, Arizona 85721 USA -
Aquaculture Pathology Laboratory
University of Arizona
1117 East Lowell Street
Building 90, Room 106
Tucson, Arizona 85721 USA -
Aquaculture Pathology Laboratory
University of Arizona
1117 East Lowell Street
Building 90, Room 106
Tucson, Arizona 85721 USA -

Aquaculture Pathology Laboratory
University of Arizona
1117 East Lowell Street
Building 90, Room 106
Tucson, Arizona 85721 USA
Related Posts

Health & Welfare
Biosecurity principles for sustainable production using SPF shrimp
Basic components of biosecurity include knowledge of diseases, adequate detection methods and the use of “clean” shrimp stocks.

Health & Welfare
A holistic management approach to EMS
Early Mortality Syndrome has devastated farmed shrimp in Asia and Latin America. With better understanding of the pathogen and the development and improvement of novel strategies, shrimp farmers are now able to better manage the disease.

Health & Welfare
A study of Zoea-2 Syndrome in hatcheries in India, part 1
Indian shrimp hatcheries have experienced larval mortality in the zoea-2 stage, with molt deterioration and resulting in heavy mortality. Authors investigated the problem holistically.

Health & Welfare
A study of Zoea-2 Syndrome in hatcheries in India, part 2
Indian shrimp hatcheries have experienced larval mortality in the zoea-2 stage, with molt deterioration and resulting in heavy mortality. Authors considered biotic and abiotic factors. Part 2 describes results of their study.