Experiences in shrimp genetic improvement in seven countries
Selection to improve growth is easy, but difficult for resistance and tolerance

The fundamental mathematical equations of quantitative genetic theory, including the one predicting selection Response, were established in the first half of the 20th century, over 80 years ago, are probably eternal and apply to all animal and plant species, and even to prokaryotes.
Under the same unchanged environment, genetics is pure and precise mathematics, an exact science that can predict everything. Introduce multiple environmental changes, especially in the extensive systems of production, and the precise mathematics of genetics becomes a very inexact science, often with little predictability.
RS is the selection response that we want to determine in our improved populations, and is based on the equation RS = (i * h * σa). RS is a factor of: i, the intensity or pressure of selection (a function of the percentage of the extreme animals we select; the lower the percentage, the greater the genetic gain); of h, which is the accuracy of the selection criteria adopted (the correlation between what we measure and the true genetic value of the animal for what we really want); and of σa, which is a function of the additive genetic variability existing in the population.
Genetic improvement: growth rate is easy, resistance and tolerance are difficult
Selecting for and achieving genetic improvements for growth is or can be easy, as there is a phenotype (weight or size) that we can measure rigorously. There is usually considerable additive genetic variability for this trait, and the correlation between the weight or size we measure and the true genetic value of the animal for what we want, growth, is high, usually > 0.6. The application of genetics in real commercial production systems is also a matter of people and human management. By combining genetic knowledge with adequate human management, and adapting them to the production and operational systems that exist, achieving genetic improvements for growth is feasible and relatively easy.

However, for survival/resistance/tolerance/general robustness, things are more complex and less straightforward. I only know of three concrete examples of successful selection for resistance/disease tolerance in vannamei shrimp, one of which is the natural selection model that has been historically used in Ecuador. The problem is that the simplest and most easily accessible phenotype we have to measure, survival, is imprecise, of a binomial statistical nature, and with a very low correlation with the true genetic value of the animal for what we really want, overall robustness before all kinds of challenges.
What we are really looking for is a shrimp that will not die, whether from diseases like White Spot Syndrome Virus (WSSV), Infectious Myonecrosis Virus (IMNV), Early Mortality Syndrome (EMS) and vibriosis; oxygen depletions, fluctuations in alkalinity and salinity; and all possible combinations of stressors.
We want a robust animal that can face all kinds of adverse conditions and also grows fast and with a low feed conversion. In the commercial production ponds of extensive systems with diseases, the true causes of mortality are often multiple and can be very variable from situation to situation. They are specific combinations of specific levels of stressors, which may sometimes include nutritional deficiencies, in the presence of pathogens.
The biological characteristics, and the genetic nature and architecture that confer survivability in a specific set of stressful situations, can be considerably different from those that enable survival in different situations. Thus, only having the survival phenotype is very limiting and greatly restricts what can be achieved through genetics. Other phenotypes to measure are needed, preferably quantitative and with wide variability, so that rapid and predictable genetic progress can be achieved for this type of characteristics. However, the reality is also that the more precise a phenotype is, the less general and often the less useful its applicability becomes. For example, years of successful efforts for the selection of resistant/tolerant shrimp to IMNV can become almost useless in a short time when the prevalent disease is changed to WSSV, which is the case for Brazil.

In view of all this, the inherent difficulties in the biological nature of what we want – coupled with permanent changes in environmental factors, in production systems and in current diseases – make specific selection for resistance/tolerance to diseases a difficult proposal that has resulted in several cases in large projects with substantial investments that ended with no tangible results. In this context, the genetic model of natural selection from commercial production ponds historically used in Ecuador and under a non-SPF (specific pathogen free)/APE (all pathogen exposed) sanitary status has brought together various very interesting characteristics, attributes and advantages.
The issue of sanitary status, SPF, High Health, or APE is another one that complicates genetic selection for disease resistance/tolerance, because according to some hypotheses, direct contact between pathogens and broodstock in grow-out ponds could lead to the development of natural mechanisms of resistance or tolerance that could not be obtained otherwise. The implication is that the SPF health status model (without direct contacts between pathogens and broodstock) would have significant disadvantages in this context of producing robust and resistant/disease tolerant animals. The available evidence from different countries seems to corroborate these hypotheses, that direct contact between pathogens and breeding populations enhances the natural mechanisms of disease resistance/tolerance. This complicates, even more the subject of genetic selection for disease resistance/tolerance.
New tools and capabilities for the implementation of genetics are now available at a new and higher level through genomics, with very high-density maps of SNPs (a type of genetic marker, a single-nucleotide polymorphism SNP is a variation in a single nucleotide found at a specific position in the genome) for selection directly at the gene level. The costs are high, but it is always, of course, a cost-benefit evaluation exercise.
The truth is that as long as the phenotype to be measured remains survival, or as long as production systems are not standardized to a much more rigorous and accurate level than today – for the reasons explained and because the fundamental equations of quantitative genetics remain the same – it is hardly credible that anything can change significantly, either with genomic selection or with any other type of more conventional genetics. Something similar could be mentioned with respect feed conversion rate, a very important productive and economic characteristic, but without a phenotype that can be easily measured at the individual or familiar level for genetic selection purposes.
Growth improvements predictable, survival rates less so
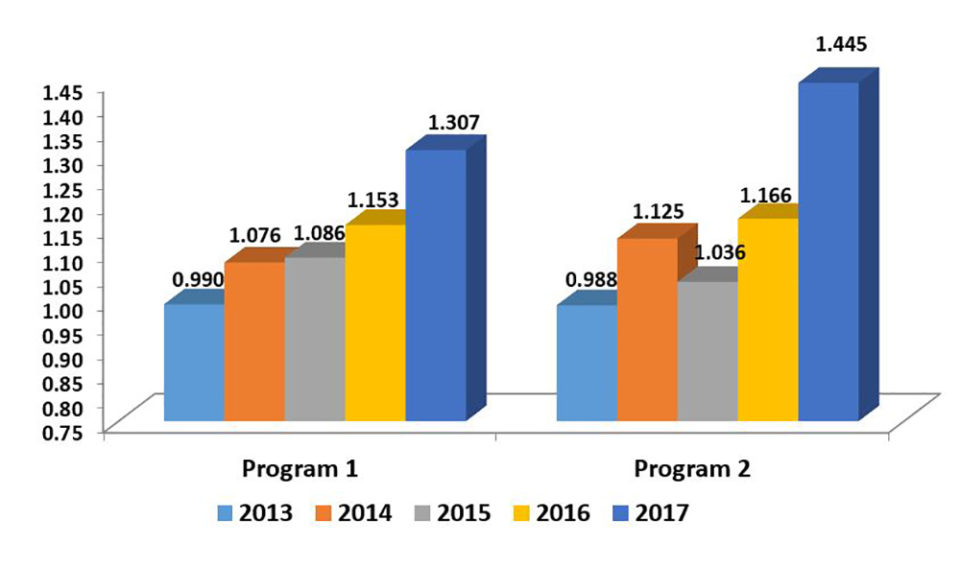
We are currently managing eight genetic improvement programs in seven different countries, one of these programs now for 14 years and the most recent one for just over a year. Growth improvements have been a constant of all programs, with annual rates between 5 and 12 percent.
The objectives, the genetic essence and methodologies, and the commercial realities of these eight programs are all different from each other. We handle all types of programs, families with elastomers, families with molecular markers, double-first cousin family systems precluding the need for physical or genetic marker identification schemes, and programs of mass selection; in addition, we have started considering genomic selection schemes (with the support of specialized companies) in some cases.
Improvements in survival rates have been less predictable and less consistent than improvements in growth rates. In general, the evolution of survival rates has been positive, but moderate and often with fluctuations that are not recorded in the most linear, predictable and constant evolution of the growth rates. In other cases, survival rates have remained fairly stable over the years without major improvements.
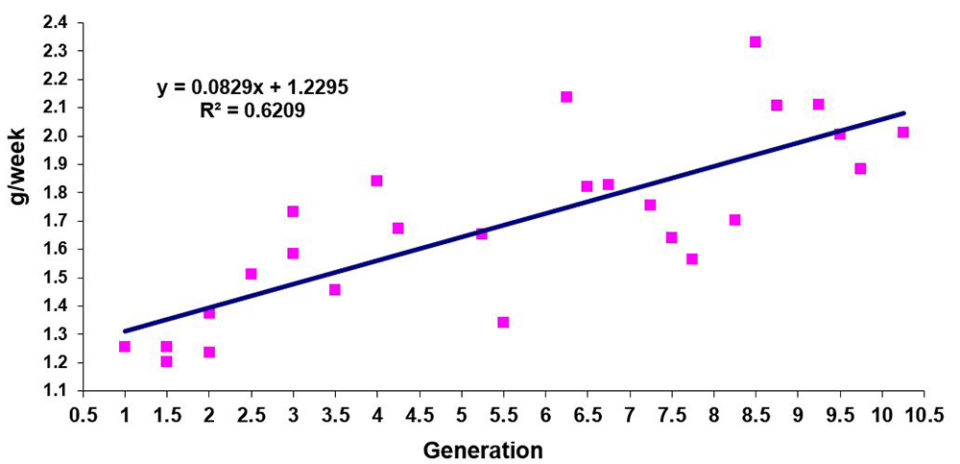
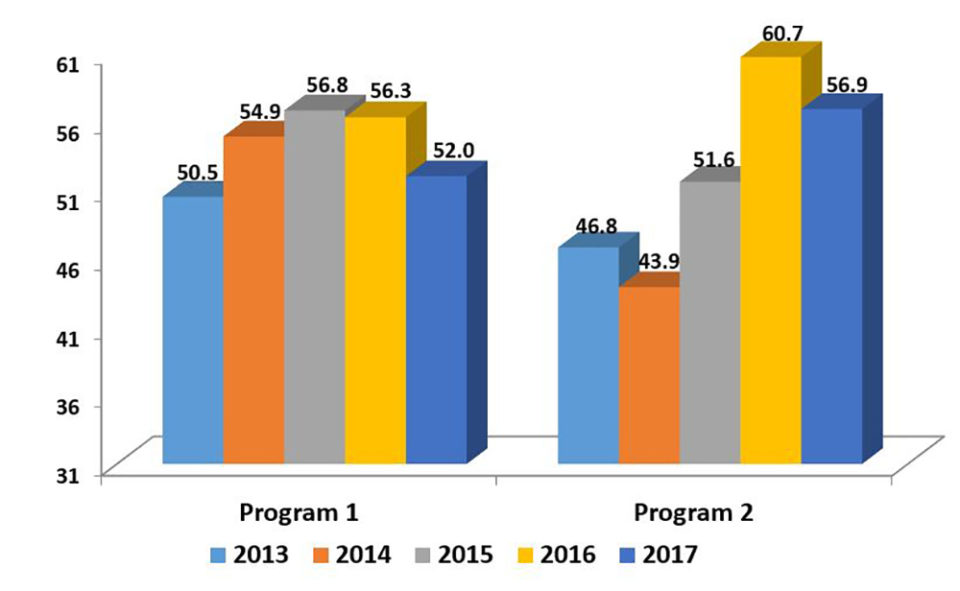

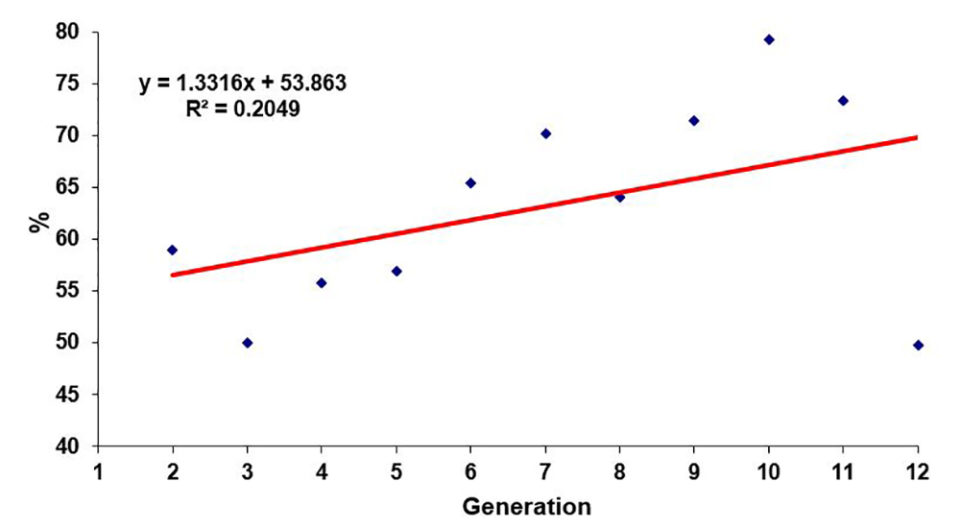
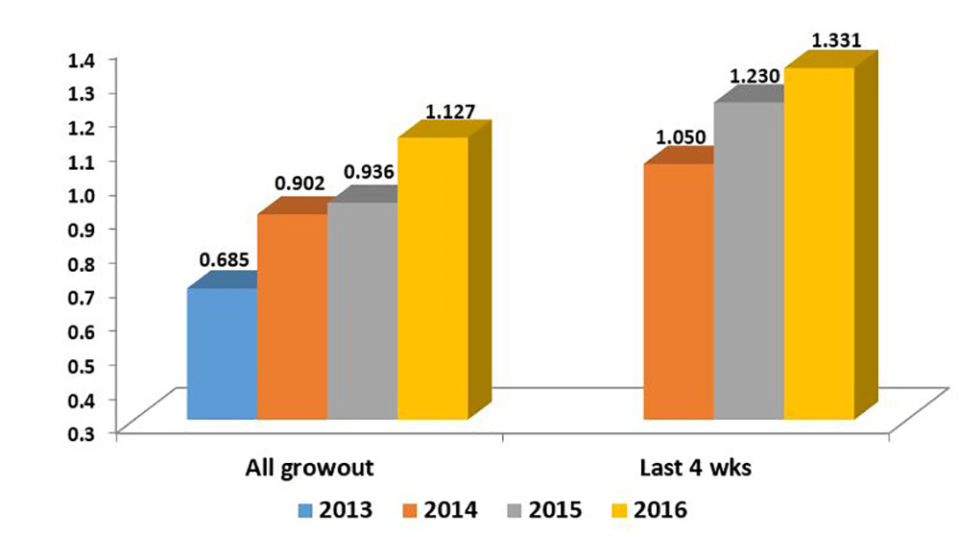
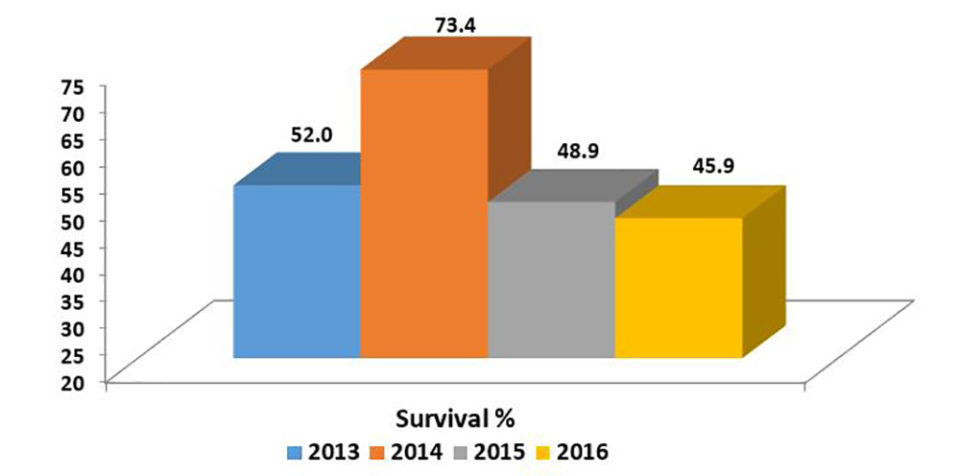
Undoubtedly, under extensive commercial conditions and especially with diseases, the correlations between growth and survival are usually negative; however, they are not of a very marked magnitude (> -0.3). For this reason, family selection programs generally allow more balanced products between growth and survival attributes. However, it is also true that, because of their nature, well-managed mass selection programs sometimes allow, in periods of a more stable climate, to mark certain growth advantages that are often quite significant.
Because of their simplicity, mass selection programs allow cost-benefit relationships that are often interesting, especially in contexts of volatility such as those that often characterize the economic reality of shrimp production. For the characteristics that most interest us in shrimp (resistance/tolerance/general robustness), realistic and feasible proposals for genomic selection schemes have been slow to appear, due to the inherent difficulties described above.

Perspectives
The emergence of new diseases in some countries of Latin America has determined situations that from a genetic point of view are very peculiar, very interesting and merit reflection. Commercial issues often prevent a frank and sincere approach to some issues at scientific meetings or in published articles. This is regrettable, because with time, a tendency is established for two different realities, the one that really happens in the aquaculture ponds, and the one that only happens in technical and scientific meetings and in published articles.
The world of shrimp genetics is not (yet) dominated by large multinational corporations, probably due to the marked level of volatility and risk in the economic reality of shrimp farming, determined primarily by new and ever-emerging diseases. This also merits some reflection, because it is not likely that it will be up to conventional genetics or to genomics to solve these issues of volatility and risk in shrimp farming. Rather, I would expect the great beneficiary will be the science of genetics when shrimp production and management systems achieve maturity.
Author
-

Consultant, Iowa Genetics
João Pessoa, Paraíba, Brazil
Related Posts

Health & Welfare
Genetics key to maximum growth rate for shrimp
Realizing higher growth rates in commercially cultured shrimp has many important benefits, including reducing various risks, cutting costs and increasing economic opportunities. Shrimp genetics primarily determines the amount of additional growth that can be achieved, as shrimp typically reflect their parents’ growth rates.

Health & Welfare
Biosecurity practices on fish farms need beefing up
Biosecurity measures and preventive strategies are essential in any biological production chain. Properly planned and implemented biosecurity programs will enhance animal health, production and economics.

Health & Welfare
Selection for disease resistance in Indian white shrimp
A new breeding program for genetic improvement of Indian white shrimp (Fenneropenaeus indicus) in Egypt was established at a private shrimp farm at AL Dibah Triangle Zone (DTZ) to develop disease-resistant shrimp for the country’s aquaculture industry.

Health & Welfare
Born in Hawaii, SPF broodstock shrimp industry faces globalization
The next step for shrimp breeding will be developing animals that aren’t just disease-free, but increasingly resistant to multiple pathogens. The industry is globalizing, with suppliers setting up shop overseas. But its birthplace will always be Hawaii.